
Introdução
Tyrannosaurus rex, Osborn 1905 (Dinosauria: Theropoda: Tyrannosauridae) foi uma espécie carnívora e bípede que viveu no fim do período Cretáceo, predominantemente na região que é hoje a América do Norte. Quando o primeiro fóssil de T. rex foi descoberto, em 1903 na América do Norte, a espécie foi logo considerada um feroz predador devido a presença de dentes enormes, pontiagudos e serrilhados dos quais, aliados às garras afiadas ficou explicito que só podiam pertencer a um matador. Entretanto, assim como é extremamente difícil estabelecer o comportamento de um animal apenas pelo seu esqueleto, garras afiadas e grandes presas nem sempre são indicativos de comportamento predatório. Os ursos contemporâneos apresentam longos caninos e garras, no entanto obtém apenas de 10% a 15 % de sua alimentação abatendo outros animais, em sua maioria de pequeno porte, com exceção da espécie Ursus maritimus, o urso polar que, devido as condições do habitat onde vive, o animal é capaz de abater caribus, bois almiscarados jovens e pequenas baleias.
Há muito que se debate se um grande teropode como o T. rex seria um predador ou um necrófago obrigatório. Em 2003, foi publicado um trabalho de Ruxton e Houston onde estes investigadores sugeriram que um enorme teropode poderia ter sido exclusivamente necrófago, já que, através da comparação com o que se passa atualmente nas savanas africanas, concluíram que haveria cadáveres suficientes para alimentá-los. Segundo o estudo, nestes ecossistemas a quantidade de herbívoros que vão a óbito diariamente é equivalente a 4,4 kg de carne por km². Se o T. rex tivesse um peso de 6 toneladas e um metabolismo reptiliano, esta quantidade de carne seria suficiente para a sua sobrevivência, desde que passasse 12 horas diárias patrulhando o habitat e se conseguisse detectar indivíduos em estado de decomposição a pelo menos 80 metros de distância. E se esse teropode conseguisse cheirar a carne dos animais mortos a 320 metros (o que alguns répteis modernos conseguem fazer), encontraria nas suas rondas diárias 3 ou 4 indivíduos, possibilitando mesmo a alimentação para um grupo formado por um número reduzido de espécimes adultos.
Ruxton e Houston (2003) não sugeriram que o seu estudo prove que um T. rex seria um necrófago obrigatório; o que concluíram é que poderia existir um número de cadáveres suficiente, de forma que o animal não fosse obrigatoriamente um predador.
Justificativa
Embora seja evidência circunstancial, Coe (1978) assinalou que as carcaças dos grandes elefantes são rapidamente liquefeitas (2 a 4 dias), através de decomposição bacteriana. O grande defensor de T. rex como um necrófago obrigatório tem sido Horner (1984, 1994). Este investigador, juntamente com outros colegas (Horner e Lessem 1993; Horner e Dobb 1997), tem apresentado vários argumentos, essencialmente relacionados com características morfológicas, no sentido de demonstrar que um estilo de vida predatório teria sido impossível para os indivíduos da espécie:
1. Olho relativamente diminuto, impedindo uma visão das presas à distância.
2. Membros posteriores com proporções sugerindo uma limitada capacidade de corrida.
3. Membros anteriores desproporcionalmente curtos, impedindo o animal de agarrar uma presa.
4. Dentes relativamente alargados, longe da configuração em forma de lâmina, típica da ordem Teropoda.
5. Grandes bolbos olfativos, que permitiriam a detecção do cheiro de carcaças a grande distância.
No entanto, muitos investigadores têm refutado estes argumentos. Farlow e Holtz (2002) verificaram que “a órbita de T. rex, em relação às dimensões do crânio, é bastante grande para um réptil da sua dimensão. Para, além disso, as dimensões da órbita sugerem que a espécie teria um olho grande em termos absolutos”. A dimensão do olho, (estimada a partir do diâmetro anteroposterior da órbita) é tão grande ou maior, em relação ao comprimento do crânio, do que em Deinosuchus, Sarcosuchus e outros teropodes de grandes dimensões. Embora vários investigadores estejam de acordo em que as proporções do membro posterior seriam menos cursoriais do que as de teropodes de menores dimensões (Christiansen 1999; Farlow et al. 2000), implicando uma capacidade de corrida relativamente reduzida (Hutchinson e Garcia 2002), Holtz (1995) verificou que as razões entre os comprimentos de metatarsos / fêmur e tíbia / fêmur de T. rex sugerem que este grande teropode “seria tão ágil ou até mais rápido do que outros grandes teropodes” (Farlow e Holtz 2002). Aliás, Hutchinson e Garcia (2002) sugeriram também que Tyrannosaurus rex, mesmo que lento, seria mais rápido do que os grandes dinossauros herbívoros contemporâneos. Sellers e Paul (2004), utilizando robótica, sugerem que um grande Tyrannosaurus rex (peso estimado em 5700 kg) poderia atingir “facilmente” velocidades superiores a 36 km / h. Relativamente aos argumentos 3 e 4 de Horner, Farlow e Holtz (2002) salientaram que vários predadores contemporâneos não utilizam os membros anteriores para agarrarem e despedaçarem as presas (“o que seria uma novidade para lobos, seriemas e aves secretário”), nem possuem dentes em forma de lâmina para matarem as presas (“o que orcas e crocodilos achariam surpreendente”).
Brochu (2000), apesar de concordar que as grandes proporções dos lobos olfativos de T. rex poderem ter sido empregues pelo teropode na detecção de carcaças em decomposição, sugeriu também que um olfato muito apurado poderia ser utilizado na detecção de presas vivas (ou para diversos comportamentos intraespecíficos).
Paul (1988) sugeriu que estes teropodes caçariam por emboscada vítimas desprevenidas; para Carpenter (2000), as presas preferenciais de Tyrannosaurus poderiam ser animais jovens e/ou doentes. “Em resumo, suspeitamos que Tyrannosaurus rex e outros teropodes carnívoros seriam, como muitos predadores atuais, carnívoros oportunistas, procurando carcaças (numa atividade em que as grandes dimensões seriam uma vantagem, Farlow 1994), mas também e sempre que possível, matando presas” (Farlow e Holtz 2002).
A ocorrência de lobos olfativos “proporcionalmente” grandes em exemplares adultos de Tyrannosaurus rex tem levado alguns investigadores a sugerirem que estes teropodes seriam essencialmente necrófagos, por comparação com o que acontece com os grandes necrófagos modernos, como os abutres. Mas, e também por comparação, não sabemos o que ocorria com outros clades e com teropodes de diferentes dimensões. “Por exemplo, a dimensão dos lobos olfativos dos tiranossaurídeos pode ser simplesmente alométrica: pode acontecer que se acabe por verificar que isto ocorre em TODOS os grandes teropodes. Enquanto não forem analisados mais teropodes gigantescos (Acrocanthosaurus, Carcharodontosaurus, etc.), será difícil afirmá-lo” (Holtz 2004). E Carpenter (2004) salientou que os resultados obtidos (ainda não publicados) para o ornitopode Camptosaurus indicam que os seus lobos olfativos eram também proporcionalmente grandes. “Para o que é que um ornitopode teria um sentido do olfato tão bem desenvolvido?; mas obviamente existe mais do que apenas localizar carne putrefata.”
Carpólitos e parte do trato intestinal de tiranossaurídeos podem fornecer algumas pistas em relação a este assunto, já que neles foram encontrados ossos de ornistiquianos juvenis (Chin et al. 1998; Varricchio 2001). Embora estas descobertas não provem que os animais eram predadores, parece confirmarem que eles consumiriam preferencialmente presas juvenis, com menos riscos e menor gasto de energia.
Meers (2003), num trabalho com o título de “A força de mordida e as dimensões das presas de Tyrannosaurus rex e as suas inter-relações para inferências do comportamento alimentar”, concluiu também que este enorme teropode do Cretácico final deveria ser um predador ativo sempre que a ocasião se proporcionava, conseguindo um animal solitário adulto atacar e matar herbívoros como Triceratops horridus, que alcançaria mais de 5 toneladas de peso (Dodson 1998), ceratopsiano muito abundante nos ambientes frequentados pelo predador.
Stevens (1998) sugeriu, através da utilização de lasers em réplicas de crânios de vários teropodes, que estes carnívoros possuíam uma visão binocular, com ambos os olhos trabalhando conjuntamente e fornecendo uma percepção em profundidade, características presentes em muitos dos predadores modernos. Verificou que Tyrannosaurus, Velociraptor e Troodon teriam campos binoculares de visão muito amplos, 50 º ou mais, semelhantes aos dos atuais falcões. Allosaurus e Carcharodontosaurus teriam um campo de visão inferior (cerca de metade), comparável ao dos atuais crocodilianos, que são “predadores de emboscada”. Stevens suspeita que a capacidade de visão global de tiranossaurídeos, velociraptoranos e troodontídeos se situaria entre a dos atuais predadores por emboscada e os predadores com maior capacidade visual. Os que possuíam campos binoculares mais amplos poderiam ter utilizado a sua estereopsia em ataques noturnos. Stevens sugere ainda que um Tyrannosaurus rex seria um predador porque os teropodes com campo de visão mais amplo também apresentam algumas características craniais, como focinhos mais baixos e uma menor quantidade de obstruções ósseas em torno dos olhos (que diminuiriam a sua capacidade binocular). Stevens sugere que a capacidade de enxergar em profundidade, juntamente com várias características do crânio, permitiriam que um teropode como Tyrannosaurus rex pudesse ter utilizado técnicas de caçada idênticas às de predadores atuais como os falcões. “É óbvio que estes dinossauros estavam bem adaptados para este estilo de vida”.

Evidências de Canibalismo
Tyrannosaurus rex foi um dos maiores carnívoros terrestres de todos os tempos sendo o quarto maior teropode registrado até então, ficando atrás das espécies (fig. A.1) Spinosaurus aegyptiacus, Carcharodontosaurus saharicus e Giganotosaurus carolinii.
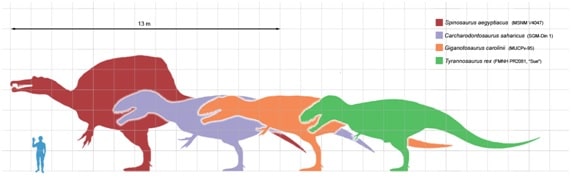
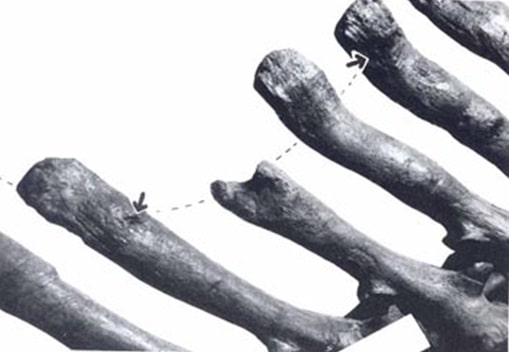
Consequentemente a ecologia e dieta de T. rex têm sido o foco de muita discussão. No entanto, há pouca evidência direta de dieta ou hábitos alimentares nesta espécie. Exames em quatro exemplares (fig. A.2) de coleções de museus da espécie revelaram marcas de dentes feitas por grandes dinossauros carnívoros, atribuídas a indivíduos da própria espécie não deixando muita dúvida nessa afirmação por conta do T. rex ter sino o único grande carnívoro a predominar a região onde hoje é a América do Norte a partir do Maastrichtiano final. As marcas são interpretadas como traços de alimentação e esses fósseis, portanto, são registros de canibalismo. Dado que este comportamento tem um potencial de preservação de baixa, o canibalismo parece ter sido uma surpresa em Tyrannosaurus, e este comportamento pode ter sido relativamente comum em dinossauros carnívoros.

Conclusão
Gould (1980) colocou uma questão pertinente: “O que um Tyrannosaurus rex conseguia fazer com os seus reduzidos braços, se é que eles serviam mesmo para alguma coisa?” Várias hipóteses foram surgindo para explicar a ocorrência, num teropode enorme, de membros anteriores proporcionalmente tão diminutos: auxílio durante a copulação (Osborn 1906), ajudar o animal a levantar-se do solo (Norman 1970); Paul (1988) sugeriu que se tratava de órgãos vestigiais, enquanto Mattison e Griffin (1989) propuseram que representaria um órgão cuja função diminuiria com a maturidade do animal.
Carpenter e Smith (2001), analisando novo material de T. rex, entretanto descoberto, concluíram que “embora proporcionalmente o membro anterior seja muito curto, a vantagem mecânica revela um sistema de forças eficiente, utilizado para agarrar as presas durante a predação”. Estes investigadores verificaram também que o músculo bíceps seria extremamente poderoso, o que associado com a presença de um úmero muito espesso e colunar, tornava os membros anteriores, não apêndices inúteis, mas utilizados com eficiência para “segurar e agarrar presas que se debatiam, impedindo que escapassem, enquanto os dentes despedaçavam o animal”. “A nossa análise da osteologia, miologia e biomecânica do membro anterior de T. rex mostra que o braço tinha uma construção robusta e era bem musculado”. Carpenter e Smith (2001) concluem que “T. rex era, portanto um predador ativo e não um mero necrófago, como tem sido sugerido”.
Um esqueleto quase completo e articulado de um hadrossauro adulto – Edmontosaurus annectens – (fig. A.3) proveniente do Cretácico final da Formação Hell Creek (Montana) fornece evidência de um ataque, de que não terá resultado a morte da presa, por parte de “um grande teropode, que se acredita poder ter sido um Tyrannosaurus” (Carpenter 2000). Neste exemplar, com cerca de 2,25 metros de altura de anca, observa-se uma seção mutilada da série caudal, em que ocorreu um «recrescimento», que Carpenter sugere poder indicar: que o hadrossauro conseguiu sobreviver ao ataque; que este ataque foi realizado quando a presa estava viva e não como um ato de necrofagia.
O topo da série caudal deste hadrossauro, se mantivesse corpo e cauda horizontalmente (Galton 1970) estaria a cerca de 2,9 metros do solo. “O único teropode conhecido da Formação Hell Creek suficientemente alto para poder ter provocado essa lesão é Tyrannosaurus rex” (Carpenter 2000). O herbívoro terá sido atacado por detrás e pelo lado direito, como é sugerido pela direção do corte efetuado na espinha neural 15. Uma grande calosidade na base do ílio indica uma fratura cicatrizada, mas extensa. Esta fratura deve ser anterior ao ataque e pode explicar a escolha deste indivíduo particular pelo tiranossaurídeo: “A função do músculo ílio-femoralis estaria comprometida e o hadrossauro provavelmente coxeava”… e “um hadrossauro coxo seria uma presa ideal para um Tyrannosaurus rex caçador”. Carpenter (2000) conclui que “Tyrannosaurus poderia ser um caçador ativo e atacar espécies presa”.
Na sua análise, Molnar (2000) conclui também que “a partir da osteologia cranial, não existe qualquer indicação que Tyrannosaurus rex não pudesse ter sido um predador ativo”. Neste estudo, Molnar conclui que as maxilas de T. rex poderiam abrir muito, o suficiente para agarrar o pescoço da maioria dos maiores hadrossauros contemporâneos. Assim, o predador poderia matar estas presas, atacando a partir de cima, quer por asfixia, quer impedindo o fluxo de sangue à cabeça, quer partindo a medula espinal. Ataques deste tipo ao pescoço de ceratopsianos serão mais improváveis, tendo em conta o enorme ‘colarinho’ parietal – squamosal destes herbívoros. Uma estratégia possível seria a de abocanhar a boca e narinas das presas, matando-as por estrangulamento, já que a boca de T. rex é suficientemente grande para agarrar o focinho e maxilas de um destes herbívoros ou de um hadrossauro.
Referências
- https://en.wikipedia.org/wiki/Tyrannosaurus
- http://dml.cmnh.org/2004Dec/msg00098.html
- http://www.dinodata.info/index.php/topic,2608.0.html


